انتقالات غشای سلولی
مقدمه
سهولت نسبتا زیادی که برای نفود مواد محلول در محیط خارج به سلول وجود دارد، معمولا با تراوایی سلولی بیان میکنند. اهمیت موضوع بسیار زیاد است. تراوایی سلول اعمال تغذیه و دفع ، تبادلات مواد بین سلولی و محیط خارج را شامل میشوند. دو لایه لیپیدی عاری از پروتئین ، اساسا نسبت به مولکولهای بزرگ قطبی و یونها ناتراوا هستند و نقل و انتقال این ترکیبات از خلال غشا در جهت یا خلاف جهت شیب الکتروشیمیایی بوسیله پروتئینهای غشا انجام میشود که نقش کانال ، پمپ یا حامل غشایی را عهدهدار هستند.
انتقال از خلال غشا غیر فعال یا فعال است. انتقال غیر فعال نوعی انتشار است که طی آن یک یون یا مولکول در جهت شیب الکتروشیمیایی و یا غلظتش حرکت میکند. در این نوع انتقال انرژی مصرف نمیشود. این نوع انتقال به دو صورت انتشار ساده و انتشار تسهیل شده صورت میگیرد. در انتقال فعال از انرژی جهت انتقال یون یا مولکول در جهت عکس شیب الکتروشیمیایی آن استفاده میشود. شیب یونی در انجام بسیاری از فرایندهای زیست شناختی اهمیت دارد. مثلا تمرکز شیبهای ترانس ممبران یونهای سدیم و پتاسیم برای هدایت تحریکهای الکتریکی در اکسون یک سلول عصبی ضروری است.
 |
انتشار ساده
در نوعی انتقال که انتشار ساده نامیده میشود مستقل از پروتئینهای غشایی است. مولکولهای هیدروفوب (آب گریز) کوچک نظیر بنزن ، اکسیژن (O2) ، نیتروژن (N2) و دیاکسید کربن (CO2) به آسانی از خلال دولایهای لیپیدی غشایی در جهت شیب غلظت عبور میکنند. علاوه بر این مواد مولکولهایی نظیر آب ، گلیسرول و اوره نیز آزادانه تحت تاثیر شیب غلظت از خلال دو لایهای لیپیدی عبور میکنند. یکی از مهمترین عواملی که سرعت حرکت ماده از طریق چربی دولایه را تعیین میکند، قابلیت حلالیت آن ماده در چربی است.
انتشار تسهیل شده
انتشار تسهیل شده یا انتقال غیرفعال نوعی انتقال در جهت شیب غلظت شیمیایی یا شیب غلظت الکتروشیمیایی (در مورد یونها و مولکولهای باردار) است که بوسیله پروتئینهای کانال یا پروتئینهای حامل غشایی انجام میگیرد. کانالهای غشایی پروتئینهای داخلی غشایی هستند که امکان انتقال سریع مولکولها و یون را از خلال غشا در جهت شیب غلظت شیمیایی یا الکتروشیمیایی فراهم میکنند. سرعت انتقال از خلال کانال حدودا 107 یون در ثانیه است.
بطور کلی کانالهای پروتئینی برای مولکولها و یونهایی که از آنها عبور میکنند، انتخابی عمل میکنند. این خصوصیت در مورد نوعی کانال غشایی که منفذ نامیده میشود، وجود ندارد. کانال و منفذ غشایی از نظر میزان اختصاصی بودن برای ماده انتقالی با همدیگر فرق دارند. جریان انتقال مواد از خلال کانال غشایی بوسیله کنترل باز و بسته شدن دریچه کانال انجام میگیرد. از این منظر کانالها را به چند گروه تقسیم میکنند.
 |
- کانال سدیم: این کانال به تغییرات ولتاژ حساس است.
- گروهی دیگر بوسیله ترکیب شیمیایی باز و بسته میشوند.
- عدهای نظیر کانال یون کلرید بوسیله AMP حلقوی گشوده میشوند.
- برخی از کانالها هم حساس به فشار بوده و با تغییرات فشار باز و بسته میشوند.
پروتئینهای حامل
پروتئینهای حامل مانند آنزیمهای متصل شده به غشا و همانند پدیده اتصال آنزیم به سوبسترا عمل میکنند. هر پروتئین ناقل دارای یک یا چند نقطه اتصالی اختصاصی است. وقتی تمام این نقاط اشباع شد، سرعت انتقال ماکزیمم است. برخی پروتئینهای ناقل میتوانند همانند آنزیم توسط مهارکنندگان ، مهار شوند که به آنها مهار کنندههای رقابتی میگویند و بعضی دیگر مهار کنندگان غیر رقابتی نامیده میشوند. پروتئینهای ناقل مواد به سه نوع میتوانند عمل کنند.
اول اینکه پروتئینهای ناقل ، ماده را فقط از یک جهت به جهت دیگر حمل کنند. دوم اینکه حمل و نقل ماده بستگی به ماده دیگری دارد ولی جهت هر دو در یک طرف است (انتقال سیمپورت) مثل انتقال گلوکز و سدیم. در سومین نوع حمل و نقل ماده در یک جهت باعث انتقال ماده دیگری در جهت مخالف آن میشود (انتقال متقابل) یا آنتی پورت) مثل پمپ سدیم _ پتاسیم.
 |
انتقال فعال
در این فرآیند ذرات در خلاف جهت شیب غلظت شیمیایی انتقال مییابند. این نوع انتقال به دو دسته انتقال فعال اولیه و انتقال فعال ثانویه که از حرکت حرکت حاملین بار در جهت شیب پتانسیل الکتروشمیایی تامین میشود، تقسیم بندی میشوند.
انتقال جمعی یا گروهی
در انتقال فعال جمعی یا گروهی که منبع انرژی آن را ترکیب پرانرژی فسفو انول پیرووات (PEP) تامین میکند، ماده منتقلشونده ضمن انتقال متحمل تغییرات شیمیایی میشود. انتقال قند گلوکز به داخل سیتوزول باکتری Ecoli و جذب آمینو اسیدها بوسیله برخی سلولهای پستانداران تابع این مکانیزم است. در غشای باکتری Ecoli پروتئین گلوکز _ فسفو ترانسفراز وجود دارد که این پروتئین ناقل گلوکز میباشد و به ازای هیدرولیز یک مولکول فسفرانول پیرووات یک مولکول گلوکز را فسفوریله میکند و همزمان به سیتوزول سلول باکتری میکند.
یونوفورها
یونوفورها خانوادهای از آنتیبیوتیکهای باکتریایی هستند که انتقال یونهای معدنی یکظرفیتی و دوظرفیتی را از خلال غشای لیپیدی زیستی و مصنوعی تسهیل میکنند. اینها مولکولهای هیدروفوب (آب گریز) کوچکی هستند که در دو لایه لیپیدی حل میشوند و غشا را به یونهای معدنی نفوذپذیر میسازند. ترکیبات یونوفور به دو خانواده حامل یونی متحرک و یونوفور کانالساز تقسیم میشوند.
انتقال فعال غشای سلولی
در فرآیند انتقال فعال ذرات در خلاف جهت شیب غلظت شیمیایی یا الکتروشیمیایی منتقل میشوند. این نوع انتقال وابسته به منبع انرژی است و منبع انرژی انتقال فعال عمدتا از ATP ، حرکت الکترونها در جهت شیب پتانسیل الکتریکی ، حرکت یونها در جهت شیب الکتروشیمیایی ، نور و ترکیب پرانرژی فسفو انول پیرووات تامین میشود.
مقدمه
یکی از اعمال مهم غشای سلولی این است که ترکیب یونی بسیار متفاوتی در داخل سلول نسبت به خارج آن ایجاد نماید. به عنوان مثال میزان سدیم خون در مهره داران و بی مهرگان 10 یا 20 برابر بیش از مقدار آن در درون سلول است و میزان پتاسیم برعکس بوده و در درون سلول 20 تا 40 برابر بیش از خارج سلول است. حفظ و نگهداری چنین شیبهایی در دو طرف غشای نیمه تراوا نیاز به مصرف مقدار زیادی انرژی دارد. انتقال از خلال غشا به صورت فعال است یا غیر فعال. در انتقال فعال از انرژی جهت انتقال یون یا مولکول در جهت عکس شیب الکتروشیمیایی استفاده میشود.
انتقال فعال اولیه
منبع انرژی انتقال فعال اولیه ATP ، جریان الکترون در جهت شیب پتانسیل الکتریکی یا نور است. برای مثال پمپ سدیم_پتاسیم و پمپ کلسیم که در انتقال فعال این یونها از خلال غشا شرکت دارند، از ATP به عنوان منبع انرژی استفاده میکنند. همچنین کمپلکسهای VI , III , I زنجیره انتقال الکترون میتوکندری ، پمپ پروتون هستند و یونهای +H را در خلاف جهت شیب الکتروشیمیایی انتقال میدهند.
پمپ سدیم _ پتاسیم
این پمپ که در غشای پلاسمایی سلولها بویژه سلولهای بافتهای تحریکپذیر نظیر بافتهای ماهیچهای و عصبی به وفور یافت میشود، در انتقال فعال آنتی پورت (در خلاف جهت هم) دو کاتیون +Na و +K نقش دارد. این پمپ به ازای انتقال کاتیونهای +Na و +K در خلاف جهت شیب الکتروشیمیایی ATP را به ADP+Pi هیدرولیز میکند.
پمپ سدیم _ پتاسیم به ازای هر مولکول ATP که هیدرولیز میکند، سه یون +Na را در خلاف جهت شیب الکتروشیمیایی از سلول خارج و دو +K را در خلاف جهت شیب الکتروشیمیایی وارد سلول میکند. از اینرو ، این پمپ یک پمپ الکتروژنیک است. حدود 10درصد اختلاف پتانسیل بین دو سوی غشا از عملکرد این پمپ ناشی میشود. بهعلاوه این پمپ نقش مستقیمی در تنظیم حجم سلول و تنظیم فشار اسمزی داخل سلول ایفا میکند.
پمپ کلسیم
یون کلسیم بهعنوان یک پیک ثانویه شناخته میشود که در انتقال پیام در داخل سلول نقش ایفا میکند. غلظت این یون در داخل سیتوزول حدود 10 درصد میکرومولار و غلظت خارج سلولی آن ده هزار برابر این مقدار است. در داخل سلول کلسیم درون اندامکهای شبکه آندوپلاسمی (ER) و میتوکندری ذخیره میشود. زمانیکه کانالهای کلسیم موجود در غشای پلاسمایی و غشای ER و میتوکندری باز میشوند، غلظت سیتوزولی کلسیم بطور آنی و به مقدار زیادی افزایش مییابد. در غشای پلاسمایی و غشای شبکه آندوپلاسمی ، پمپهای کلسیمی وجود دارد. اینها با مصرف ATP به منظور کاهش مجدد غلظت کلسیم سیتوزولی این یون را به خارج سلول یا به درون لومن شبکه آندوپلاسمی پمپ میکنند.
انتقال فعال ثانویه
منبع انرژی انتقال ثانویه از حرکت یونها در جهت شیب پتانسیل الکتروشمیایی تامین میشود. در سلولهای پوششی جدار روده و سلولهای توبولینی کلیه ، پروتئین انتقال دهندهای وجود دارد که بطور سیمپورت (همجهت باهم) گلوکز و کاتیون +Na را به درون سلول منتقل میکند. این پروتئین یون +Na را در جهت شیب الکتروشیمیایی و گلوکز را در جهت شیب شیمیایی داخل سلول منتقل میکند. به عبارتی این پمپ بخشی از انرژی که از انتقال یونهای +Na در جهت شیب الکتروشیمیایی حاصل میشود را صرف انتقال مولکول گلوکز در خلاف جهت شیب شیمیایی میکند.
 |
|
|
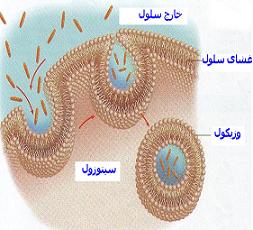
آندوسیتوز
اکثر مواد از طریق انتشار یا انتقال فعال به درون سلول راه مییابد. اما ذرات بسیار بزرگ بواسطه یک عمل تخصص یافته غشای سلولی موسوم به آندوسیتوز وارد سلول میشود. اشکال اصلی آندوسیتوز عبارتند از پینوسیتوز (قطره خواری) و فاگوسیتوز (ذره خواری).
پینوسیتوز
غشای اکثر سلولهای دائما در حال پینوسیتوز میباشد اما این کار در برخی سلولها سریعتر است مثلا در ماکروفاژها. پینوسیتوز تنها راه ورود اکثر مولکولهای درشت به درون سلول میباشد. به عنوان مثال خلاصه پینوسیتوز مولکولهای پروتئینی را شرح میدهیم. این مولکولها معمولا بر گیرندههای تخصص یافته بر روی سطح غشا متصل میشود که مخصوص پروتئینی هستند که باید جذب شود. بطور کلی گیرندههای در گودههایی کوچک به نام گودههای پوشیده در سطح بیرونی غشای سلول متراگم شدهاند.
در زیر گودهها و بر روی سطح درونی غشای سلول ، شبکهای از یک پروتئین فیبریلی به نام کلاترین و نیز پروتئیهای دیگر وجود دارد که احتمالا شامل فیلمانهای انقباضی اکتین و میوزین میباشد. هنگامی که مولکولهای پروتئینی به گیرندهها متصل میشود خصوصیات سطح غشا چنان تغییر میکند که تمام گوده به درون سلول فرو میرود و پروتئینهای پیرامون آن (اکتین و میوزین) تدریجا به هم نزدیک میشوند و لبههای گوده به هم میرسند. بلافاصله پس از آن ، قسمت فرو رفته غشا از سطح سلول جدا میشود و یک وزیکول پینوسیتوزی در سیتوپلاسم سلول بوجود میآید. انرژی این عمل از مولکول ATP تامین میشود و ضمنا برای این عمل وجود یون کلسیم در مایع خارج سلولی ضرورت دارد.
فاگوسیتوز
شبیه پینوسیتوز است با این تفاوت که بجای مولکولها با ذرات بزرگ سر و کار دارد. تنها برخی سلولها معین قادر به فاگوسیتوز هستند از جمله ماکروفاژهای بافتی و برخی گلبولهای سفید خون. هنگامی که پروتئینها با پلی ساکاریدهای واقع بر سطح ذراتی همچون باکتری ، سلول مرده یا سایر بقایای بافتی به گیرندههای رو ی فاگوسیت متصل میشوند، فاگوسیتوز آغاز میشود.
در مورد باکتریها ، معمولا باکتری قبلا به این آنتیبادی خاصی متصل شده است و اتصال آنتیبادی به گیرندههای فاگوسیت موجب کشیده شدن هر دوی آنها به درون فاگوسیت میشود. به این عمل واسطهای آنتیبادیها اپسونیزاسیون گویند.
اگزوسیتوز
خروج مواد بزرگ یا مولکولهای درشت از سلول و ورود آن به مایع خارج سلولی توسط پدیده اگزوسیتوز انجام میگیرد. معمولا باقیمانده مواد هضم شده توسط لیزوزومها که دیگر قابل تجزیه نیستند و به جسم باقیمانده معروف هستند توسط عمل اگزوسیتوز از سلول خارج میشود. عمل اگزوسیتوز درست عکس عمل اندوسیتوز میباشد.